
Amphidomataceae
by Koyo Kuwata, Mitsunori Iwataki, Mona Hoppenrath & Urban Tillmann (December 2025)
Below you will find general morphological characters that can be used for identification of the taxa of the family Amphidomataceae, the genera Amphidoma and Azadinium and its species.
Their so far known molecular phylogenetic relationships are illustrated in this tree.

General light microscopy appearance
Most species of Amphidomataceae are small (10–20 µm in length), with both varieties of Azadinium caudatum and a few species of Amphidoma being notable exceptions. Most species share an ovoid to elliptical cell shape and may exhibit variable size ratios between the hypo- and epitheca. Under light microscopy (Fig. 1), the parietally positioned plastid is visible, as is the deep and wide cingulum, which accounts for roughly one-fifth to one-quarter of the cell length, along with a very distinct, pointed cell apex with the apical pore complex (APC). Additional features observable under light microscopy includes the large nucleus and, in some species, an antapical spine — although in certain species the spine may be very small and difficult to detect— and, where present, one or (in some species) multiple pyrenoids, which are recognizable by their characteristic starch caps.
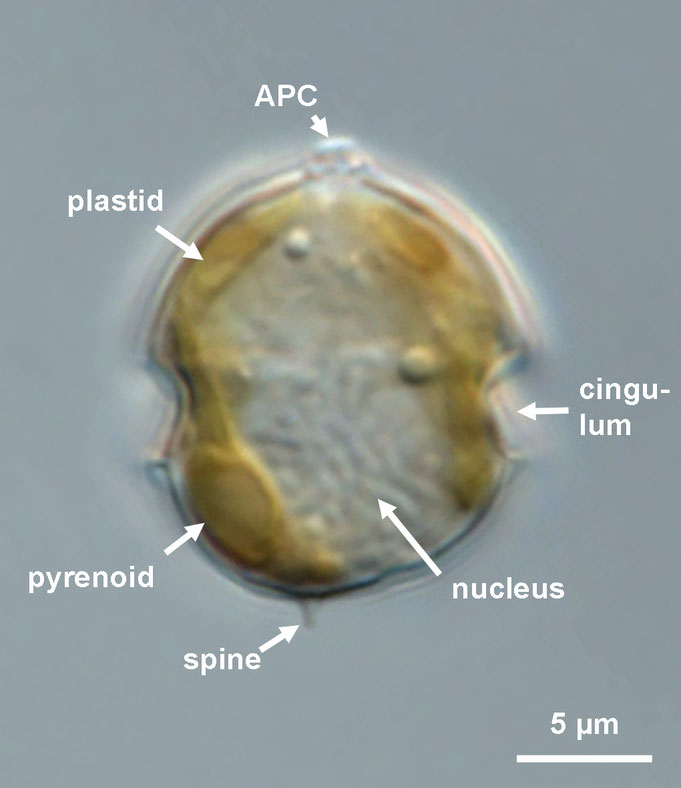
Fig. 1: Amphidomataceae, general light microscopy appearance.
Swimming pattern
All Amphidomataceae species have been reported to display a distinctive swimming behavior. Typically, cells move at relatively low speeds — for example, one strain of A. poporum swims at approximately 400 µm s⁻¹, while for Azadinium caudatum var. margalefii the speed has been recorded as 85 µm s⁻¹. The swimming paths are irregularily interrupted by brief, rapid “jumps” in various directions. Some Amphidoma species are particularly slow in their overall movement, as e.g. reflected in the epithet of Amphidoma languida (from the Latin languida, meaning lazy, slow). High-speed jumps occur intermittently but are consistently observed, especially when cells encounter nearby objects, such as the glass bottom of the observation chamber. Occasionally, cells may cover larger distances at elevated speeds, although such events are infrequent.
video clips coming soon ...!
Plastid, pyrenoid, nucleus
All species of Amphidomataceae are photosynthetic and, as demonstrated for A. spinosum, are capable of growth in axenic culture. Each species possesses a presumably single, parietally arranged chloroplast that is lobed and extends into both the epi- and hypotheca (Fig. 2A-E). In several species, a stalked pyrenoid is visible under light microscopy due to its characteristic starch cup (Fig. 2F-M), and the presence, absence, or position of the pyrenoid provides a useful feature to support species-level differentiation within the Amphidomataceae. The nucleus is thick with multiple chromosomes, and exhibits species-specific shapes ranging from round to ellipsoidal, or rarely elongated, with slight variability in its position within the cell (Fig.2F-P).
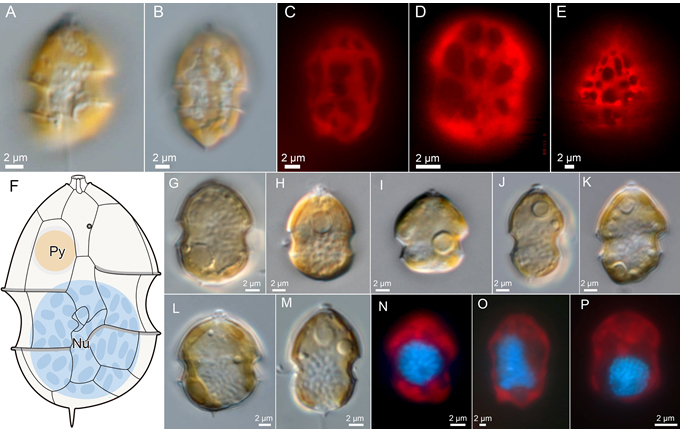
Fig. 2: Plastid, pyrenoid, nucleus. C–E, and N–P: Epifluorescence to indicate the chloroplast (red) or the DAPI-stained nucleus (blue in N–P).
Ultrastructure
Transmission electron microscopy (TEM) observation of intracellular ultrastructure reveals general features of dinoflagellates, including the dinokaryotic nucleus (Nu), mitochondria with tubular cristae, parietal chloroplasts (Ch) surrounded by three membranes, a round to oval pyrenoid (Py), and fibrous bodies (Fv) (Fig. 3).

Fig. 3: TEM sections of three different species to illustrate general cell ultrastructure.
Three types of pyrenoids — stalked, bulged, and embedded — have been observed within the family.
Cytoplasmic invagination into the pyrenoid matrix has been observed in Amphidoma but not in Azadinium (Fig. 4, lower panel), suggesting that the presence or absence of this invagination may represent a genus-level diagnostic character.
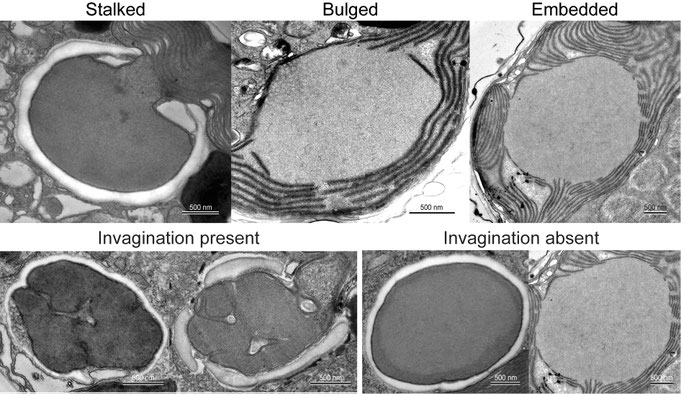
Fig. 4: Details of pyrenoid ultrastructure.
Thecal plates
Amphidomataceae possess a theca composed of cellulose plates;
however, these plates are very thin and, in most species, are difficult to discern under the light microscope. When the protoplast leaves the theca under stress (a process known as ecdysis, Fig.
5A-F), theca becomes visible even by light microscopy (Fig. 5).

Fig. 5: The theca becomes visible after ecdysis of cells. A–C and D–E: Time series of the same cell during ecdysis.
The plates can also be visualized by fluorescence microscopy after cellulose staining (Fig. 6) and examined with respect to their arrangement.
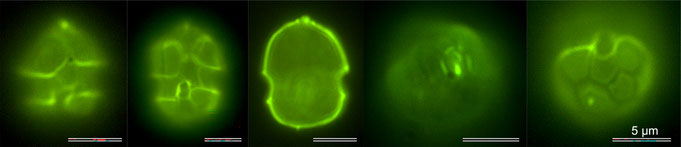
Fig. 6: Epifluorescence visualization of thecal plates after staining with Solophenylflavine.
For a complete and detailed analysis of the plate pattern and further diagnostic features of the theca, however, the use of electron microscopy (SEM, Fig. 7) is recommended.
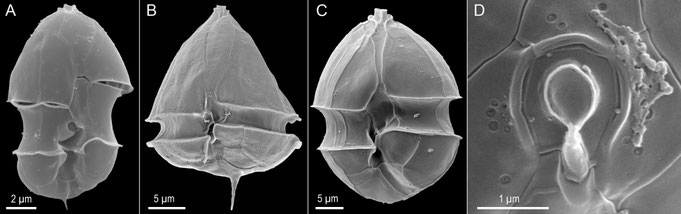
Fig. 7: Examples of SEM images for the analysis of plates and other structural details.
Plate pattern
All amphidomatacean species share the distinctive combination of six plates in each of the precingular, cingular, and postcingular plate series (Fig. 8), a feature not observed in other thecate dinoflagellates and thus representing a synapomorphy of the Amphidomataceae. Within the family, a major difference exists in the epithecal plate arrangement: all Azadinium species possess three or four apical plates and two or three anterior intercalary plates, whereas all Amphidoma species have six apical plates and lack intercalary plates entirely (Fig. 8, lower panel). All species share two antapical plates of different size as well as five sulcal plates.
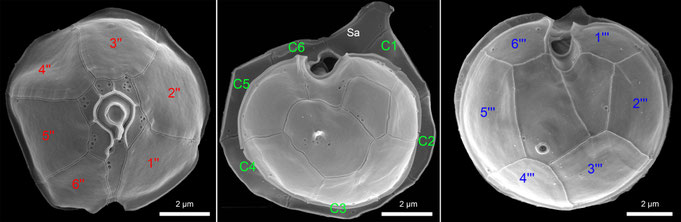
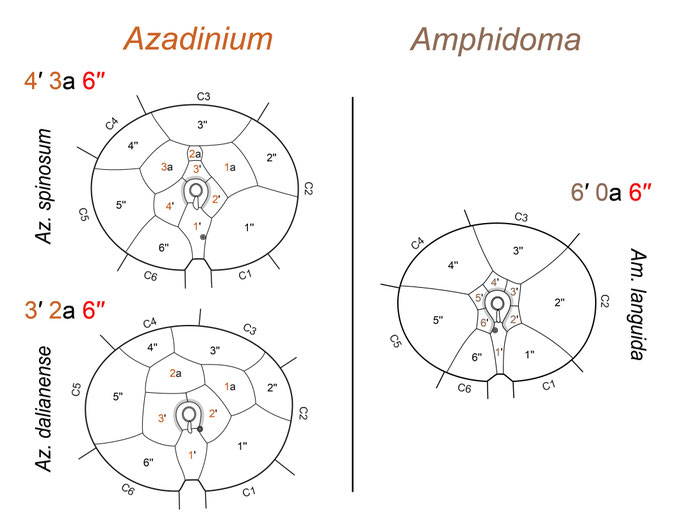
Fig. 8: The general plate pattern, shown by representative SEM micrographs (upper panel) and a schematic illustration (lower panel).
Plate surface
The surface of thecal plates in most species is smooth (Fig. 9A, B), although some species exhibit rough (Fig. 9C, D) or strongly reticulated (Fig. 9E, F) plate ornamentation. In general, the plates are perforated by small pores, which may vary in density and distribution: they can be numerous and randomly arranged, as in A. caudatum; sparse and scattered, as in smaller Azadinium species; or particularly concentrated on the apical plates in certain Amphidoma species. In some Amphidoma species, a distinct row of pores may occur below the lower cingulum (postcingular) list, and additional rows of pores can be present along the sutures between apical and precingular plates (not shown).

Fig. 9: Examples of different plate surface structures.
APC morphology
A highly characteristic feature among Amphidmataceae is the prominent apical pore complex (APC), which consists of a pore plate (Po) with a central, elongate or slightly protruding round pore covered by a cover plate (cp), and a small X-plate in contact with the pore plate and the first apical plate (Fig. 10). The pore plate varies in shape from round to slightly ellipsoidal, distinctly elongated, or markedly asymmetric, and is surrounded by a conspicuous rim (Fig. 10C) whose form differs slightly among species. The small X-plate is positioned at the junction where the Po abuts the first apical plate (1′) and occupies approximately one-third of the connection between Po and 1′ (Fig. 10C, G). Its size is most clearly observed from an internal view only (Fig. 10C), whereas externally the X-plate exhibits a consistent and distinctive three-dimensional structure in all Amphidomataceae, characterized by finger-like protrusions that connect to the apical cover plate (Fig. 10B, E-H).
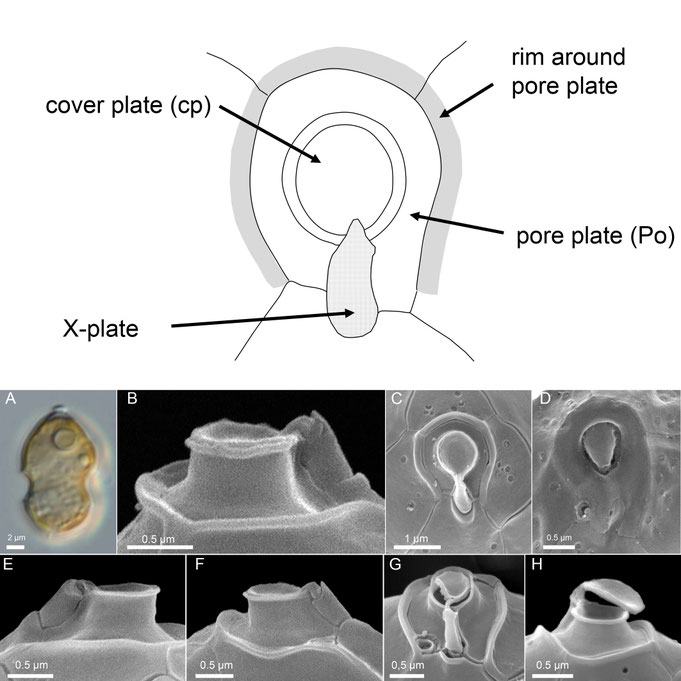
Fig. 10: Morphology of the apical pore complex (APC).
Ventral pore
With the exception of some species of Amphidoma, Amphidomataceae typically possess a distinct pore located in the epitheca (Fig. 11, white arrows), historically designated as the “ventral pore” (vp) based on its position in the first described species, Azadinium spinosum. The amphidomatacean vp is larger than typical thecal pores, surrounded by a platelet-like structure, and exhibits species-specific positions on the ventral side of the epitheca. In many species the vp is located within or in contact with the pore plate (Fig. 11D-K). The position of the ventral pore serves as a highly informative diagnostic character for species-level identification within the family.
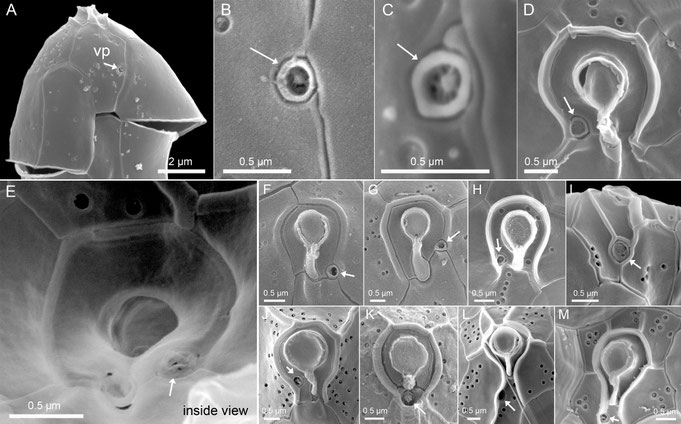
Fig. 11: Examples of the structure and position of the ventral pore (white arrows) (note that E shows an internal view).
Antapical structures
The posterior part of the hypotheca (the second antapical plate) can be smooth and without special structures in some species of Amphidomataceae, but in other species (Fig. 12) this plate bears the singular antapical spine, which can vary greatly in length and expression among species, and may also be located either at the end of a horn (Fig. 12B, C) or at the antapical end of a generally triangular, tapering hypotheca (Fig. 12D, E).

Fig. 12: Examples of a single antapical spine at the end of the hypotheca, where present.
In some cases, though not in a species-stable manner, the second antapical plate may exhibit a field of several to many small pores (Fig. 13A-C), either in addition to the spine or, in species lacking a spine, as the only structure. Observed so far only in certain species of Amphidoma, a so-called “antapical pore” may also be present here (Fig. 13D-F). This “pore” limited by a distinct rim in fact is a depressed field containing a larger number of small pores (Fig. 13E, F).
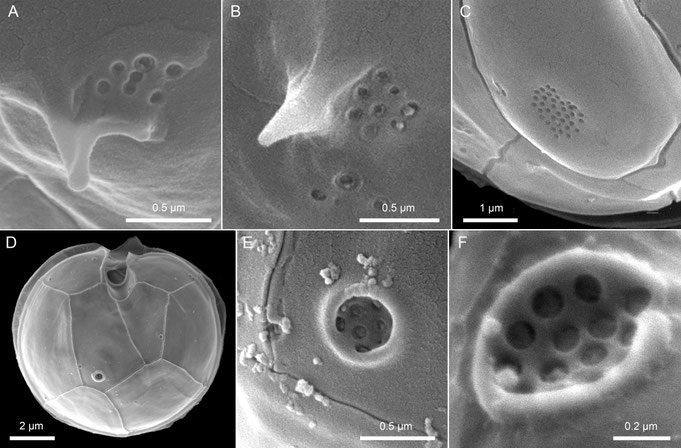
Fig. 13: Examples of potential pore arrangement on the hypotheca (A–C) and the morphology of the so-called antapical pore (D–F).
Sulcal plate arrangement
The sulcal plate arrangement is generally difficult to analyze because the pronounced three-dimensional shape of the sulcal area obscures some of the smaller plates from view. Nevertheless, all amphidomatacean species possess five sulcal plates with the same overall configuration (Fig. 14, upper panel), characterized by a large anterior sulcal plate (Sa) extending into the epitheca and a large posterior sulcal plate (Sp). Moreover, there is a distinctive and conserved left sulcal plate (Ss) which runs between and thus contacts both terminal cingular plates C1 and C6. Two small plates (Sd and Sm) form an inverted pocket where both flagella emerge, which is most clearly visible in internal view (Fig. 14). It should be noted that this sulcal plate arrangement differs markedly from that found in both the Peridiniales and the Gonyaulacales (Fig. 14, lower panel).
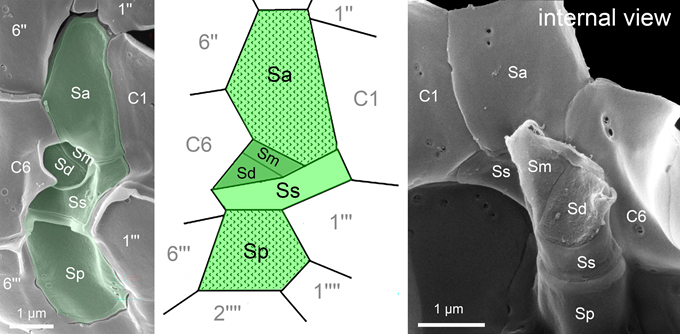
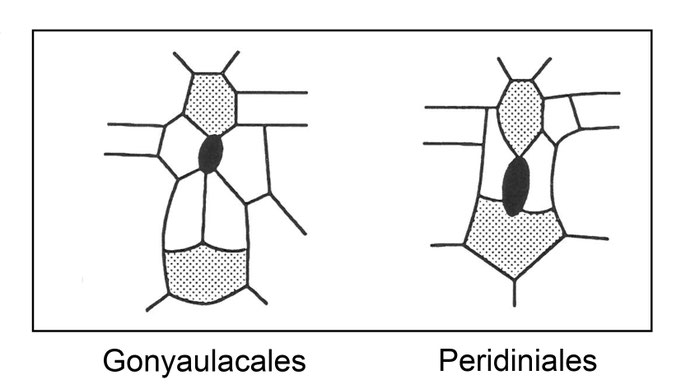
Fig. 14: Sulcal plate arrangement of Amphidomataceae (upper panel) and comparison with the arrangement of Gonyaulacales and Peridiniales (lower panel).
Cell dividion
Amphidomatacean species reproduce vegetatively through simple binary fission (Fig. 15). Dividing cells retain motility throughout both mitosis and cytokinesis. The first visible sign of mitosis is an enlargement of the nucleus, which becomes elongated and extends obliquely across almost the entire cell length. The nucleus then divides along its longitudinal axis. Cytokinesis begins slightly before nuclear division is complete (Fig. 15, upper left panel) and occurs via desmoschisis, meaning that the parent theca splits along well-defined, conserved fission lines and is then shared between the two daughter cells (Fig. 15, upper right panel). The left portion of the parent cell retains the apex, including the apical pore complex and all apical and epithecal intercalary plates, while the right portion retains, among other structures, both antapical plates.
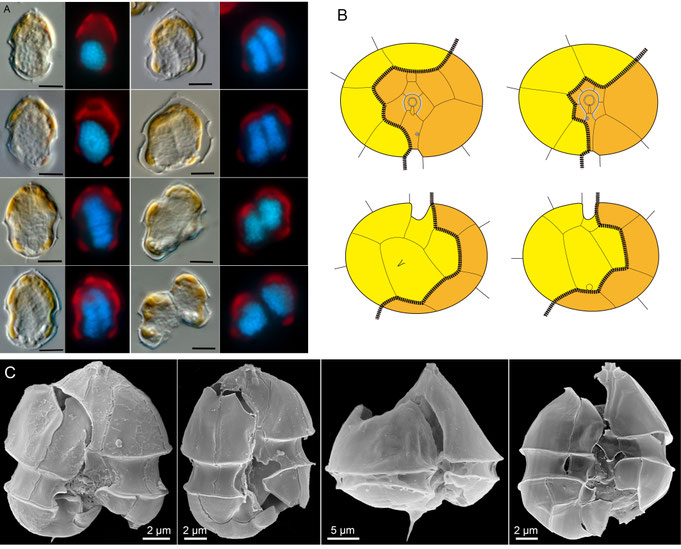
Fig. 15: Cell division in Amphidomataceae. Upper left: reconstructed light microscopy time series of cell division in different cells, each shown in bright field and after nuclear staining. Upper right: schematic representation of the fission line (black line) in Azadinium (left) and Amphidoma (right). Lower panels: representative SEM micrographs of dividing cells.
Coccoid resting stages
Knowledge of the life cycle of Azadinium and Amphidoma remains incomplete. The successful isolation of Az. poporum from incubated sediment samples strongly suggested the presence of coccoid resting stages, a finding that has since been confirmed for a few Azadinium species (Fig. 16). These coccoid stages appear only rarely and sporadically in culture, are small and round to slightly ellipsoid, and contain pale granules, sometimes accompanied by a yellow accumulation body (Fig. 16N). Early stages of these coccoid cells exhibit chlorophyll fluorescence (Fig. 16J-L), whereas mature stages show greatly reduced fluorescence and can persist for extended periods in an apparently unaltered state. This suggests that they may function as long-term survival stages rather than temporary cysts, although it remains to be determined whether these stages are part of a sexual life cycle. Scanning electron microscopy (Fig. 16O) has not yet revealed any external structures, such as paratabulation or an archeopyle, and hatching of these stages has not been observed.
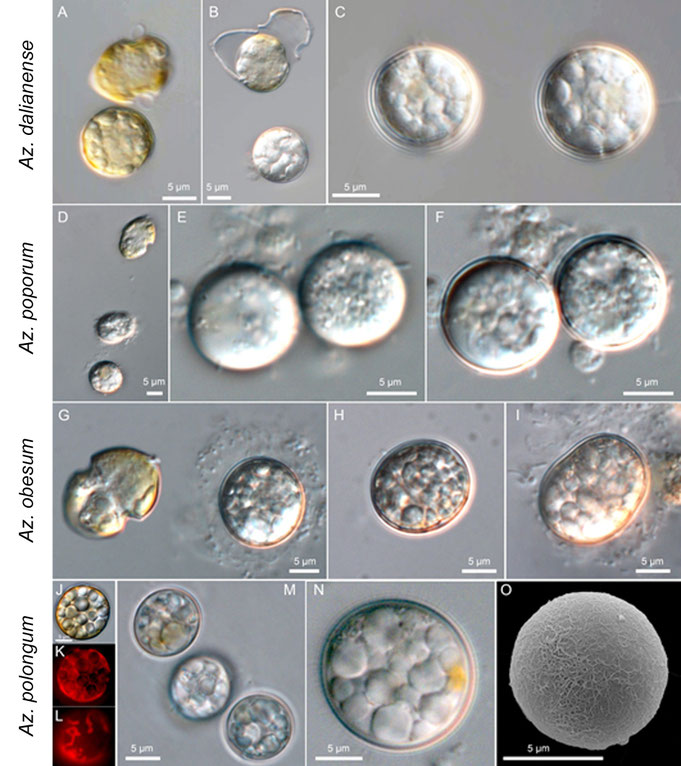
Fig. 16: Examples of coccoid resting stage formation in some species of Amphidomataceae.
SENCKENBERG am Meer
Deutsches Zentrum für Marine Biodiversitätsforschung
PD. Dr. Mona Hoppenrath
Südstrand 44, 26382 Wilhelmshaven
